
Article Categories
- All Categories
-
 Data Structure
Data Structure
-
 Networking
Networking
-
 RDBMS
RDBMS
-
 Operating System
Operating System
-
 Java
Java
-
 MS Excel
MS Excel
-
 iOS
iOS
-
 HTML
HTML
-
 CSS
CSS
-
 Android
Android
-
 Python
Python
-
 C Programming
C Programming
-
 C++
C++
-
 C#
C#
-
 MongoDB
MongoDB
-
 MySQL
MySQL
-
 Javascript
Javascript
-
 PHP
PHP
-
 Economics & Finance
Economics & Finance
Plant Growth Regulators and Their Mode of Action
Introduction
Plant growth regulators which are also called plant hormones are the ones that regulate the growth of a plant. These PGRs act as signaling molecules. These PGRs are produced within the plant in extremely low concentrations.
These hormones are produced by the cells of a plant and not all regulate the growth of the plant but also determine the formation of other parts of the plant like flowers, leaves, shedding of leaves, development, and ripening of fruits, etc. In short, PGRs control all the physiological processes within the plants.

Types of Plant Growth Regulators and their Mode of Action
The following are the main PGRs that are produced by plants.
Auxin
Auxins have an amino acid precursor, tryptophan. It is a growth-promoting hormone that facilitates cell wall loosening, increases cell walls? plasticity, and helps in cell division and expansion.
Its major role is seen in root growth promotion. NAA prevents premature leaves and fruit drops. It leads to apical dominance due to which the primary shoot grows more than compared to lateral shoot. Removal of the apical bud leads to enhanced growth of the lateral shoot. It promotes parthenocarpy due to which seedless fruits are produced. Examples-seedless strawberries and grapes.
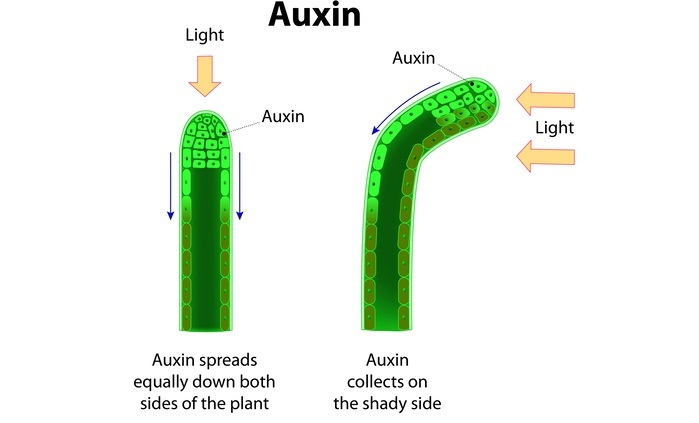
Mode of Action
Key players: Auxin acts as a ligand and TIR1 (transport inhibitor response 1) is an auxin hormone receptor. TIR1 works with other F-box proteins like SCF which is associated with SKP proteins and Qulin proteins. TIR1 protein acts as an E3 ligase that can attach polyubiquitin to the target protein for its destruction.
ARF auxin response factor is a transcription factor that helps in transcribing the genes necessary to provide a response against the auxin. AUX/IAA is an inhibitor of ARF. 26S is a proteolysis complex.
For every signaling pathway, activation of transcription factors occurs and transcription factor activates certain response genes.
In the nucleus, we have regulatory regions and structural genes that will be transcribed into target mRNA which will be utilized for producing proteins.
When the plant does not receive auxin, ARF is inhibited by AUX/IAA inhibitor and does not bind to the regulatory region. Hence no structural gene transcription is possible.
-
When auxin is present, it is first transported to the plant cell and gets bind to the receptor present in its cytoplasm. This binding activates the F Box complex as a result of which TIR1 protein gets activated and starts adding the multiple polyubiquitins into the AUX/IAA inhibitor which leads to its degradation through the 26S proteolysis complex.
Then ARF which acts as a transcription factor becomes free to bind to the regulatory region in the nucleus as a result of which the structural genes will be transcribed into specific mRNA and later get translated into proteins in the cytoplasm. These proteins are necessary for cell growth.
Gibberellin (GA)
It has isoprenoid compound precursor IPP. GA?s most important role is in internode and shoot elongation. This makes it suitable for increasing the yield of sugarcane. GA stops the seed and bud dormancy and stimulates their growth.

Mode of Action
GID1 is a gibberellin receptor and a soluble protein in nucleus and cytoplasm. First the gibberellin hormone which is produced by the seeds binds to the GID1 receptor and enhances the interaction of GID1 and DELLA proteins which are repressors that inhibit plant growth.
This forms the GA-GID1-DELLA protein complex causing the delta chain structure allowing proteins called Prefoldings to bind which then helps to degrade this protein. A transcription factor called PIF binds to the complex and transcribed the plant DNA to mRNA which expresses gibberellin response genes.
This mRNA then goes to a ribosome where it is translated into protein typically an enzyme called amylase. This enzyme then functions to break down starch into sugar, an energy that the cell or seed can use to energize plant growth and elongation.
Cytokinin
It has isoprenoid compound precursor IPP. Cytokinins in contrast to auxins prevent apical dominance and promotes lateral shoot growth. It induces cell division and also delays senescence. The most active and naturally occurring cytokinin is Zeatin.
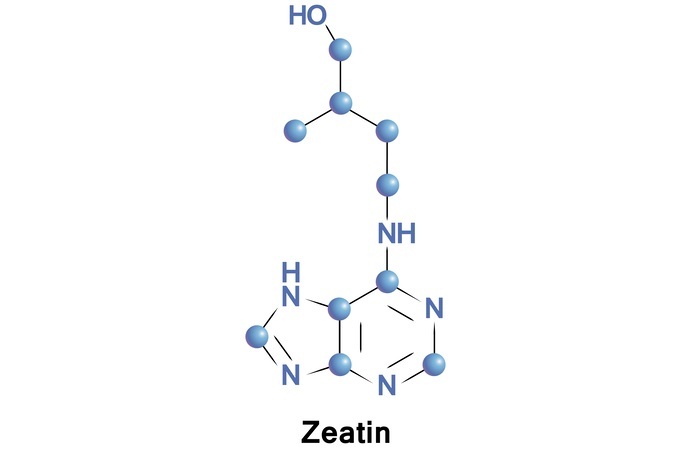
Mode of Action
This pathway is a kind of two-component regulatory system pathway. Here, cytokinin is the ligand and CRE-1 (cytokinin response-1), AHK, AHK-2, AHK-3 (Arabidopsis His kinase) are the receptors. These receptors have the CHASE domain and cytokinin binds to this domain. AHP (Arabidopsis histidine phosphotransfer protein) and ARR (Arabidopsis response regulator).
Cell membrane has 2 monomers of CRE-1 or AHK receptors. Each receptor has outer CHASE domain and inner HIS KINASE and RECEIVER domain. Now cytokinin comes and binds to the receptors at CHASE domain to drive the pathway.
This ligand binding induces the dimerization of CRE-1 or AHK receptors thereby activating the HIS kinase. The activated HIS kinase acts on ATP and gets the phosphate from it. This Phosphate is transferred to the receiver domain of the receptor. The phosphate is then transferred to the inactive AHP protein in the cytoplasm.
This AHP then moves into the nucleus and transfers the phosphate group to the aspartate residue of type B ARR. This then regulates the DNA and transcription of type A ARR-producing mRNA which later translates and produces type A ARR protein. This type A ARR receives the phosphate group from AHP.
This ARR is the one that regulates the cytokinin-responsive genes. This type A ARR forms a negative feedback loop with type B ARR thus inhibiting
Ethylene
Ethylene is also derived from an amino acid precursor, methionine. It increases the respiration rate and is well known for its role in fruit ripening and promotes senescence and root hair formation. It leads to the lateral growth of the plant.

Mode of Action
As ethylene is a negative regulator, its absence drives the pathway and its presence will stop the pathway.
Plants have various ethylene receptors which are present in the endoplasmic reticulum. All are structurally related and dimeric multi-pass proteins. These receptors have copper ions in their ethylene binding domain. These receptors also have a domain that interacts with protein CTR-1.
In the absence of a hormone, the ethylene receptor is active and this keeps CTR-1 active. The active CTR-1 has an active serine/threonine kinase domain. The function of CTR-1 in ethylene signaling depends on the activity of this kinase domain and its association with the N-terminal domain with the ethylene receptor.
The active CTR-1 then undergoes an unknown signaling mechanism as a result the EIN-3 is ubiquitilated and degraded in the proteasome. Due to this degradation, the transcription of the ethylene response gene cannot be formed.
In the presence of ethylene, it diffuses into the cell through the cell wall and into the ER and here in ER lumen it binds to the receptor. This binding inactivates the receptor, as a result, conformational changes occur and CTR-1 cannot bind to this receptor. Hence CTR-1 is inactivated and the downstream signaling pathway coming from it is blocked. In this case, EIN protein is not ubiquitilated and degraded. Now EIN-3 is translocated into the nucleus and binds and activates the transcription of ethylene target genes.
Abscisic Acid (ABA)
ABA has isoprenoid compound precursor IPP. It has a growth inhibitory role where it suppresses bud and shoot growth and often acts as a gibberellin antagonist. During various abiotic and biotic stress conditions, ABA level increases which imparts stress tolerance to the plant. It also results in stomatal closure.
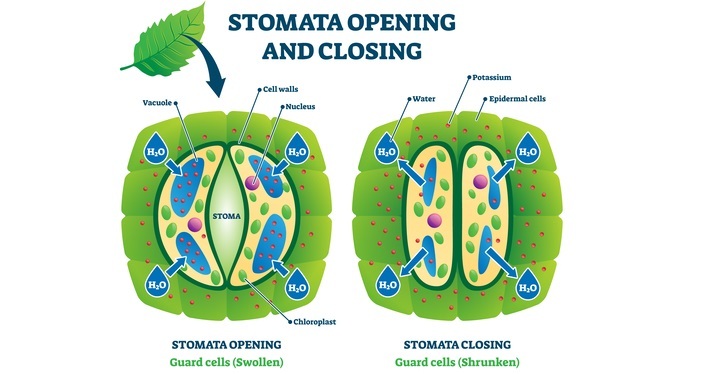
Mode of Action
ABA is a drought hormone that regulates the stomatal opening and closing. Stomata has 2 guard cells and a water pore in the center. Plants need to open stomata in the daytime in order to get the CO2 and water.
During every climate, plants need to close the stomata to avoid water evaporation. So in the presence of sunlight, there is no role of ABA hence stomata are opened. Guard cells have calcium storage vacuoles and these vacuoles have different potassium, calcium, and chloride ion channels.
Also, the guard cells have potassium (influx and efflux the potassium ions), chloride 9exports the chloride ions) ion channels and a proton pump. ABAR is a receptor found on the membrane of the guard cell and the signaling begins when there is a ligand ABA.
ABA interacts with ABAR due to which PLC (cell membrane-associated component phospholipase C) produces IP3 which interacts with the calcium channels in the vacuole and allows the calcium to be transported to the cytoplasm of guard cells leading to calcium influx (primary ca influx). ABA once activated also activates reactive oxygen species (ROS) which causes calcium influx (secondary) through calcium ion channels. Both this calcium influx causes a high concentration of calcium inside the guard cell.
On one hand, these calcium ions bind to the potassium ion influx channel and block it. On the other hand, this calcium binds to the chloride export channels and allows the export of chloride ions outside. This calcium also binds to the proton pump and inhibits its function leading to a high concentration of hydrogen ions making it acidic inside the guard cells.
Due to these potassium export channels are activated and it is balanced by the efflux of chloride and calcium ions from vacuoles. Due to the rapid movement of potassium and chloride ions outside the guard cell, an osmotic imbalance is created. Hence water moves outside the guard cells and results in the stomatal closure.
In presence of the sun, the proton pump is activated which has ATP synthase activity and transports hydrogen ions outside the guard cell. As a result of this osmotic imbalance is created. Then it activates K influx carriers and then sugars produced by the plants move inside the guard cells via the sugar transporter. Due to this imbalance, water rushes inside the guard cells. This results in stomatal opening.
Conclusion
Plant growth regulators are also known as plant hormones or phytohormones. These are the organic substances that are produced naturally in plants and involve in controlling growth and many other physiological processes within the plants moving from one part to other even though present in tiny amounts. These act like chemical messengers within the plants influencing many patterns of plant development.

1K+ Views
